今天推送的文章发表在上的“SuccessfulManipulationoftheProductSpectrumoftheErwiniaamylovoraLevansucrasebyModifyingtheResiduesaroundloop1,Loop3,andLoop4”,作者为江南大学沐万孟教授。
左旋蔗糖酶(LSs,)是属于糖苷水解酶家族68(GH68)的糖基转移酶,催化合成2,6-β-D-果聚糖。当以蔗糖为底物时,左旋糖苷酶催化不同链长的果聚糖的形成,包括聚合度20的低聚果糖,[低分子质量,FOSMW5×104Da]Levan,[高分子质量,MW5×104Da]Levan。到目前为止,已经测定了来自淀粉性埃文氏菌(EaLS,PDBID:4D47)、枯草芽孢杆菌(BsSacB,PDB:1OYG)、重氮葡萄球菌(GdLS,PDB:1W18)、巨大芽孢杆菌(BmLS,PDB:3OM2)、塔斯马尼埃文氏杆菌(EtLS,PDB:6FW)和布伦氏菌(BrsLS,PDB:7EHR)的六种LS的晶体结构。所有这些LS都显示出一个类似的五叶片β-螺旋,这在GH68和32成员中是典型的,包围了一个共同的底物结合中心腔。在这个位置,果聚糖通过由三个保守氨基酸(BsAcB中的Asp86、Asp247和Glu342)协调的双置换反应机制合成。
除了催化残基,在过去的几年里还发现了底物周围的其他残基。到目前为止,LSs已经成功地结晶成一系列的复合物,包括蔗糖(Ea-LS,PDB:4D47)、蔗糖(Bs-SacB,PDB:1PT2;Brs-LS,和PDB:7FDZ)、棉籽糖(Bs-SacB,PDB:3BYN)和Levan型FOS(Bs-SacB,PDB:6VHQ),提供了丰富的糖结合亚基信息。这些位点被发现对产品谱很重要。由于相同的双取代催化机制,所有LSs中的水解亚中心,特别是−1亚基都高度保守。除了−1亚基外,是否存在对Ea-LS产物重要的残基,以及Bs-SacB的OB位点是否会对Ea-LS中的产物分布产生类似或不同的影响,都引起了人们的兴趣。在这项研究中,对这些OB位点进行了替换突变,目的是找出哪些残基可能导致Ea-LS的产物分布,以及它们如何影响最终产物谱。结果成功地获得了一系列不同产物谱的突变体。突变体G98E、V151F和N200T均显著提高了HMWLevan的相对分子质量和产率,表明Ea-LS的聚合得到了成功的加强。
解析LSs的结构分析
到目前为止,已确定了六个LSs的晶体结构,并比较了它们的结构和序列相似性,如表1所示。结果表明,Ea-LS具有较高的Et-LS和Brs-LS的结构相似性,但与Bm-LS和Bs-SacB的结构相似性较低。Ea-LS与Et-LS的同源性最高,达90.29%,而与Bm-LS(27.75%)和Bs-SacB(28.27%)的同源性不到30%。与其他LSs相比,Ea-LS的序列相似性和结构相似性一致。

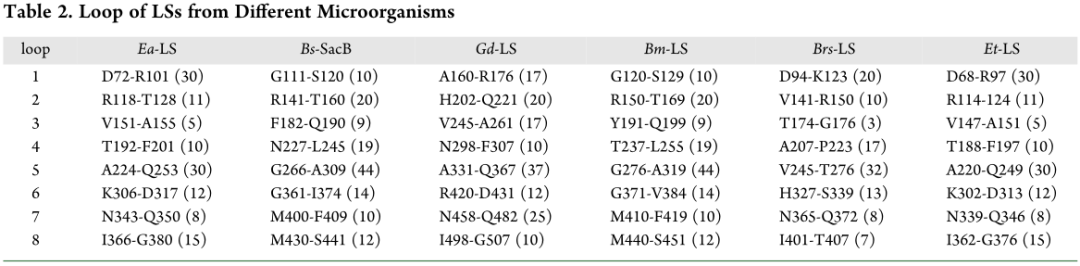
9个高变区环形成了LSs的中央口袋(图1)。Ea-LS、Bs-SacB、Bm-LS、Gd-LS、Et-LS和Brs-LS中央口袋边缘的这些环的长度和位置如表2所示。与其他LS相比,Ea-LS和Et-LS在中央腔外有一个更长的环1(30个残基)。如图1C所示,低聚糖被延伸到Bs-SacB的环3和环4。与Ea-LS和Et-LS相比,在其他LSs中,这些环更长,特别是在Bs-SacB和Bm-LS中,它们在该区域显示了几乎相同的序列。Bs-SacB的+3和+4亚位均位于环1、3和4。Ea-LS和Bs-SacB的催化口袋如图1B,D所示。Ea-LS的体积和表面积均小于Bs-SacB,中央口袋的表面积和体积与产物在LSs中的分布之间的关系尚未揭示。
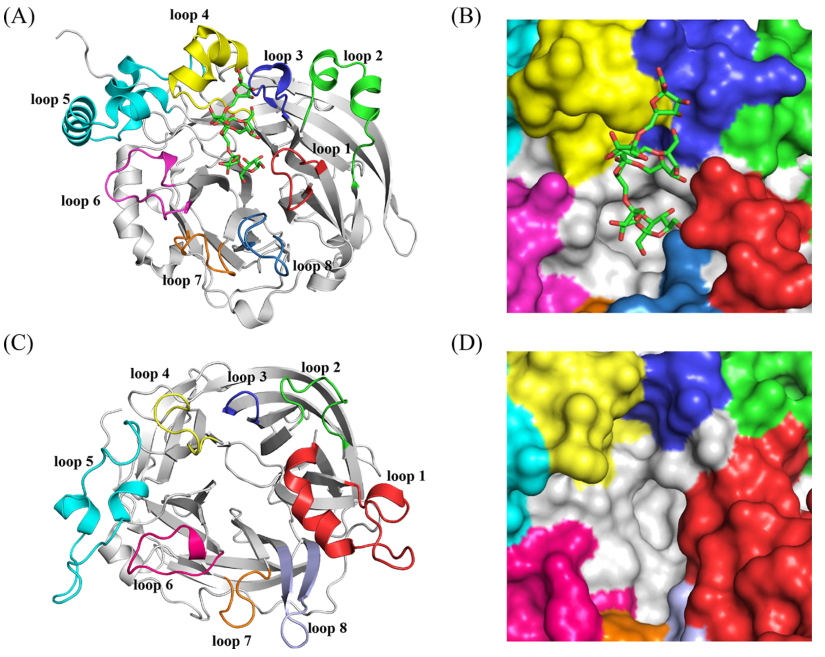
OB位点的序列比对和变异性
在这项研究中,多蛋白结构比对和用于分析解析的LS的氨基酸变异性。图2显示并比较了这些LSs中OB位点的相应残基。因此,+1亚位点的谷氨酸(BsSacB中的340)和精氨酸(BsSachB中的360)分别与谷氨酰胺(EaLS中的285)和组氨酸(EaLS中的305)半保守。Ea-LS的残基T308对应于+2亚位点BsSacB的K363,被发现保守为苏氨酸或赖氨酸。在+3亚位点中出现在BsSacB的F182对应位置的缬氨酸在革兰氏阴性细菌LS中完全保守,但在革兰氏阳性LS中不保守。此外,在革兰氏阴性和革兰氏阳性细菌LS中未表现出高度保守性的残基G98(BsSacB中的D117)、A197(Bs-SacB中的D241)、F198(Bs-SacB中的N242)和N200(Bs-SacB中的T244)也被Bs-Sac中的相应OB位点取代。
突变体对酶活性和转糖基化与水解反应比率的影响
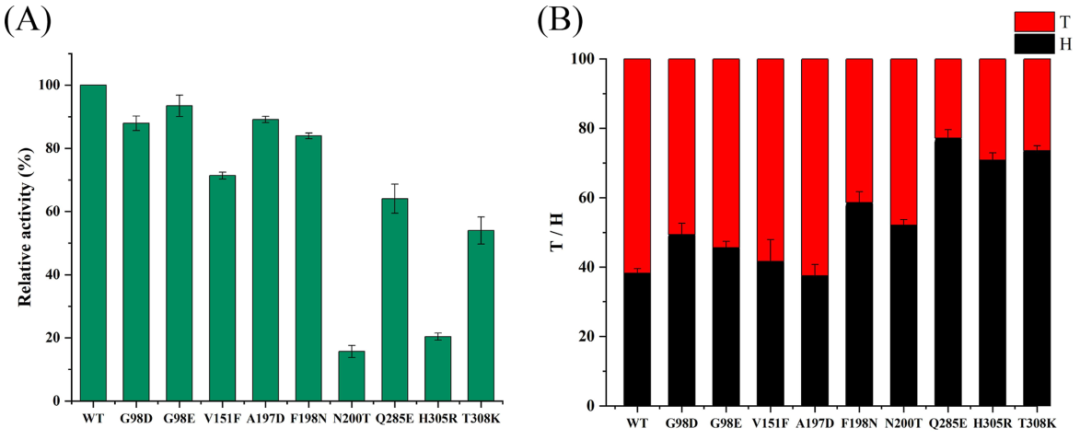
九个突变体N200T、Q285E、H305R、F198N、A197D、G98D、G98E、V151F和T308K被成功表达和纯化(图3)。WT酶和九种突变体的活性和转果糖基化-水解反应(T/H)均在相同条件下测定。结果,包括G98D、G98E、A197D和F198N在内的大多数突变体显示出与WT酶相当的活性(图4A)。V151F、Q285E和T308K等其他突变体保留了大约50-70%的酶活性。然而,突变体N200T和H305R的活性急剧下降。相比之下,移动发酵单胞菌LS(Zm-LS)的突变体H296R和H296W(EaLS中的H305)仅保留了部分活性,而同一位置的其他突变体几乎失去了活性。Brs-LS+1亚位点的突变体H327A也显示酶活性显着降低。与WT酶相比,地衣芽孢杆菌8-37-0-1LS(Bl-LS)的突变体N251A(相当于Ea-LS中的F198)仅显示出不到20%的活性。在Ea-LS中用Asn替换F198保留了大部分酶活性,表明Asn的羰基可能维持了酶与底物之间的接触。
除了突变体A197D(图4B)外,大多数突变体的T/H都有所下降。同时,突变体G98D、G98E、V151F和A197D仍以催化转果糖基化反应为主导反应。相比之下,其他突变体,包括Q285E、H305R和H308K,显示转果糖基化活性显着降低,但与WT相比,它们的水解酶活性增加。在这些等同位置的LSs报告了类似的结果。发现丁香假单胞菌pvLS(Lsc3)突变体Q301E(Ea-LS中的Q285)和D300N(Ea-LS中的D284)更喜欢水解而不是转果糖基化反应。值得注意的是,突变体D300N显示T/H降低,但与WT酶相比,它合成果聚糖而不是FOS。此外,Lsc3的突变体E110D具有更高的T/H比,并表现出高Mw果聚糖产量的减少。所有这些结果表明T/H与LSs的主要反应相关,但与最终产物谱不完全一致。
突变体对动力学参数的影响
WT酶和突变体的动力学参数如表3所示。水解和转移酶反应的Km分别表示为“KmH”和“KmT”。突变体H305R表现出KmH和KmT均增加,表明突变体H305R的蔗糖亲和力显着降低。相比之下,Q285E突变体显示出比WT酶增加的蔗糖亲和力,因为它的KmH和KmT都较低。突变体F198N显示KmH降低但KmT增加,表明与WT酶相比,它更喜欢水解蔗糖。突变体G98D、G98E和N200T都表现出与WT酶相当的KmH,而G98D和G98E的KmT增加了。
突变体对产物分布的影响
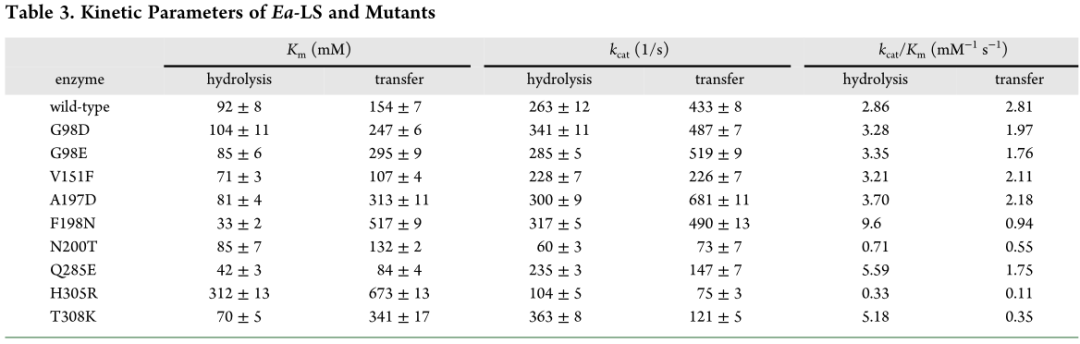
本研究中,检测到FOS和果聚糖分别使用HPAEC-PAD和HPGFC(图5A,C)。结果发现WT酶在反应混合物中合成FOS(主要是DP6)和HMW果聚糖(MW为4.4×107Da)。同时,九个Ea-LS突变体呈现出不同的产物光谱,通常可以分为三组。第1组:A197D、F198N、Q285E和T308K,显示HMW果聚糖产量降低;第2组:G98D和H305R,显示出类似的HMW果聚糖分布;第3组:G98E、V151F和N200T,其在HMW果聚糖的分子量和产率方面都表现出明显的增加。
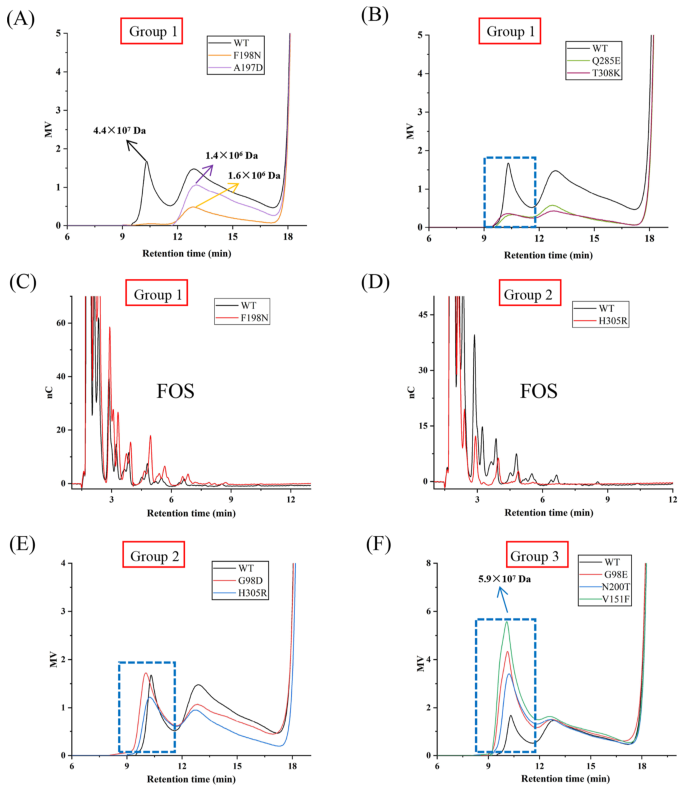
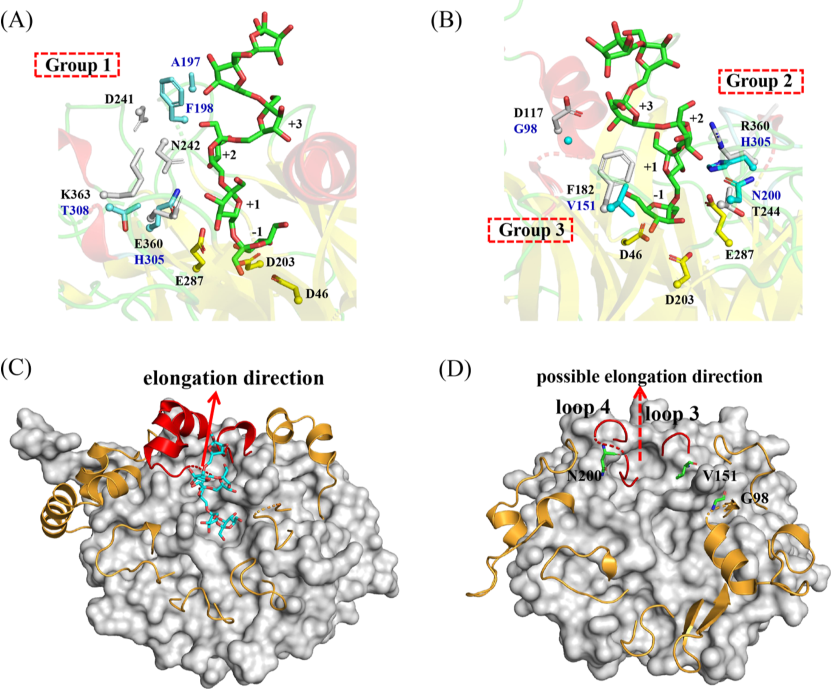
Ea-LS的第1组突变体:突变体Q285E和T308K保留了部分合成HMW果聚糖的能力,其MW与WT酶相似,但果聚糖的产量显着降低(图5A、B)。相比之下,突变体F198N和A197D不仅表现出产量下降,而且还表现出果聚糖MW的降低。谷氨酸或谷氨酰胺可在GH68酶Q285的对应位置找到,属于“DxxER”基序,是+1亚位点的一部分。在蔗糖复合物(PDB:1PT2)中,Bs-SacB中的E340(Ea-LS中的Q285)通过强氢键与葡糖基部分的3-和4-羟基接触,将果糖基和葡糖基单元锁定到确定的方向并围绕催化三联体协调它们(图6A)。该位点阻止了底物的取向,导致活性降低,尤其是聚合活性降低。类似地,Bm-LS中的突变体E350A和Lsc3中的Q301A表现出酶活性的显着降低(表4)。Lsc3中的Q301E突变体几乎完全能够分解蔗糖,但它的聚合活性降低了。
与WT酶相比,F198N突变体产生较少的果聚糖但产生更多的FOS(图5C)。与F198不同,T308远离LSs的蔗糖结合腔。尽管如此,该残基可能会稳定FOS的第三个果糖基单元,并将其引导至合适的位置,作为进一步转果糖基化的新型受体。用赖氨酸取代Ea-LS中的残基T308阻碍了其合成左聚糖的能力。关于突变体A197D,虽然其产物谱也表现出较大的差异,但目前对其潜在功能的报道有限。

Ea-LS的第3组突变体:残基G98、V151和N200预计远离催化三联体并靠近通道表面(图6D)。图6B显示了这些位点在Ea-LS中的位置和相互作用。Bs-SacB中的残基D117(Ea-LS中的G98)通过氢键参与构成+3亚位。与天冬氨酸相比,谷氨酸具有更长的侧链,这可能有利于左聚糖聚合物的延伸。这种变化也可能通过氢键在酶结构的上部形成相互作用,降低共价中间果糖基酶的分解速率,增加寡糖链接收下一个受体的时间(图6B)。
Bs-SacB中的F182(Ea-LS中的V151)通过疏水相互作用稳定寡糖链的第四个果糖基单元。同时,F182支持与W163(Ea-LS中的W131)的堆叠相互作用,这对于协调配体在亚位-1和+1处的结合至关重要。虽然这两个残基在Ea-LS中的功能尚未见报道,但相应残基V151(相当于Bs-SacB的F182)发生了突变。因此,突变体V151F在HMW果聚糖生产中表现出显着变化,表明在151位点取代苯丙氨酸可能会增强第四个果糖基的相互作用,并与Ea-LS中的W131建立堆积相互作用。
根据BsSacB的左聚糖六糖复合结构,发现左聚糖延伸方向主要受环3和环4的影响,并由一系列OB位点协调,包括D117(Ea-LS中的G98)和F182(Ea-LS中的V151)(图6C)。在Ea-LS中,突变体G98E和V151F可能具有类似的功能来稳定FOS并将其引导到环3和环4旁边。相比之下,EaLS的环3和环4比SacB的环更灵活和更短,它提供果聚糖聚合物延伸的更大空间(图6D)。事实上,G98E和V151F突变体都显示Ea-LS的HMW果聚糖产量增加。
Levan型FOS对WT和突变体的影响
作者将3%纯化的FOS(MW=2.1kDa)添加到反应中,以进一步探索突变对酶的底物和产物特异性的影响。18小时后,WT和突变体都显示LMW果聚糖产量增加,但HMW果聚糖产量没有增加(表5),表明FOS的添加增强了Ea-LS的非加工机制。Bs-SacB及其突变体S164A也观察到类似的结果,表明FOS可能是比蔗糖更好的受体(2%)。

文章信息:
DOI:10.1021/















